
Beta Diversity of Plant Communities in Relation to Soil C:N:P Stoichiometry across 150 Years of Vegetation Restoration in a Temperate Zone
 , , ,
, , ,  and
and {kind=link}
{kind=link}
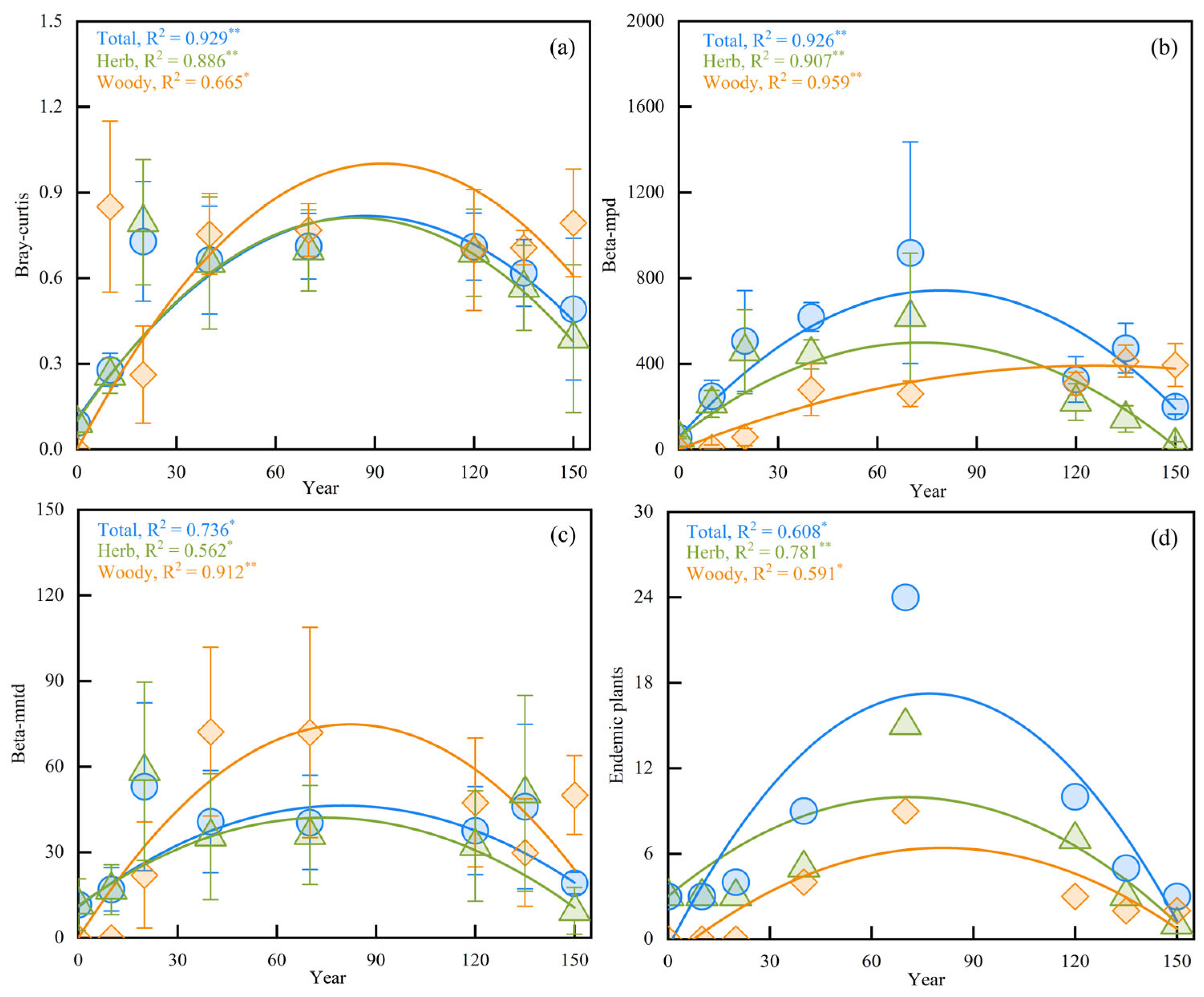{kind=link}
{kind=link}
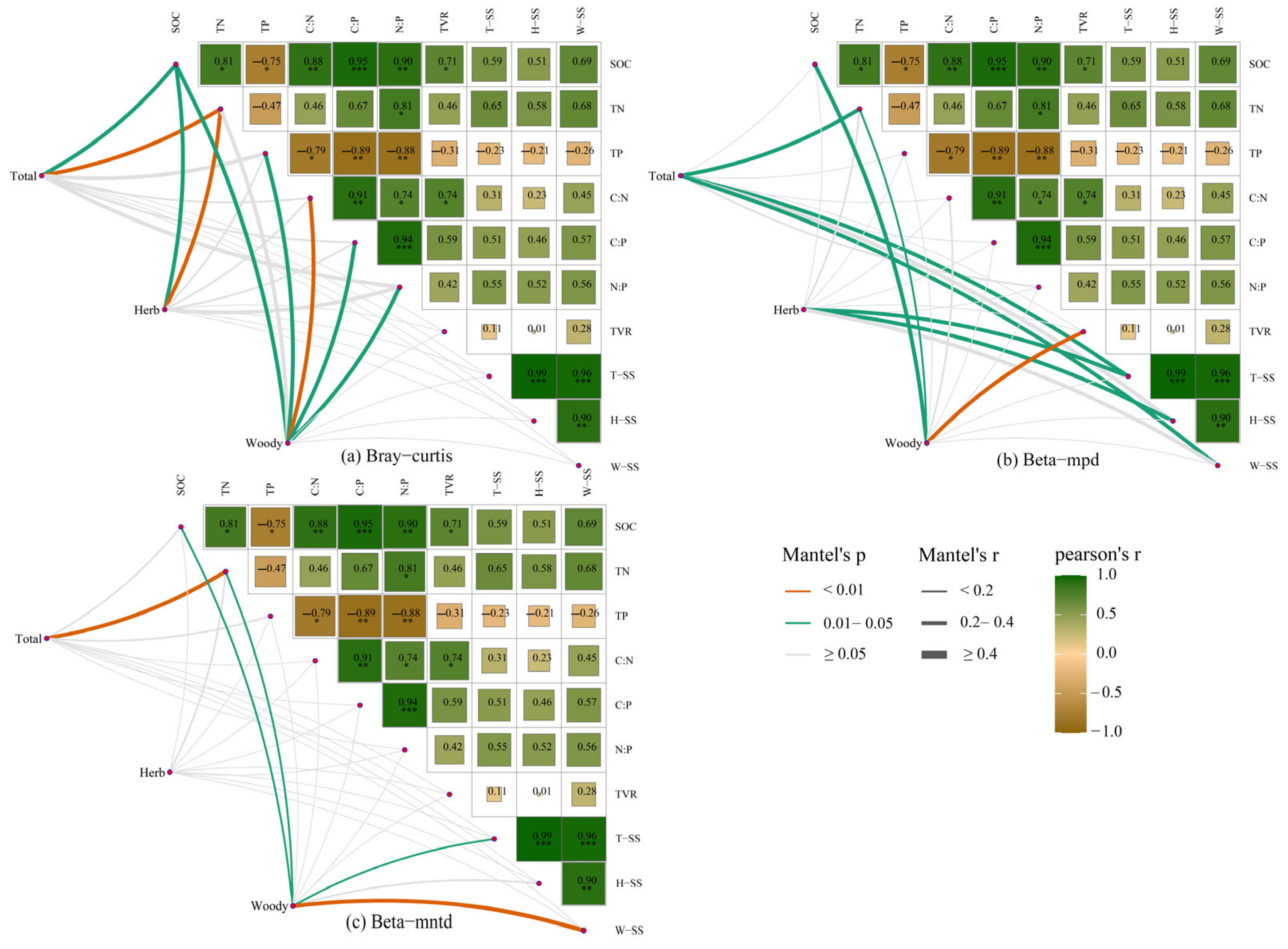{kind=link}
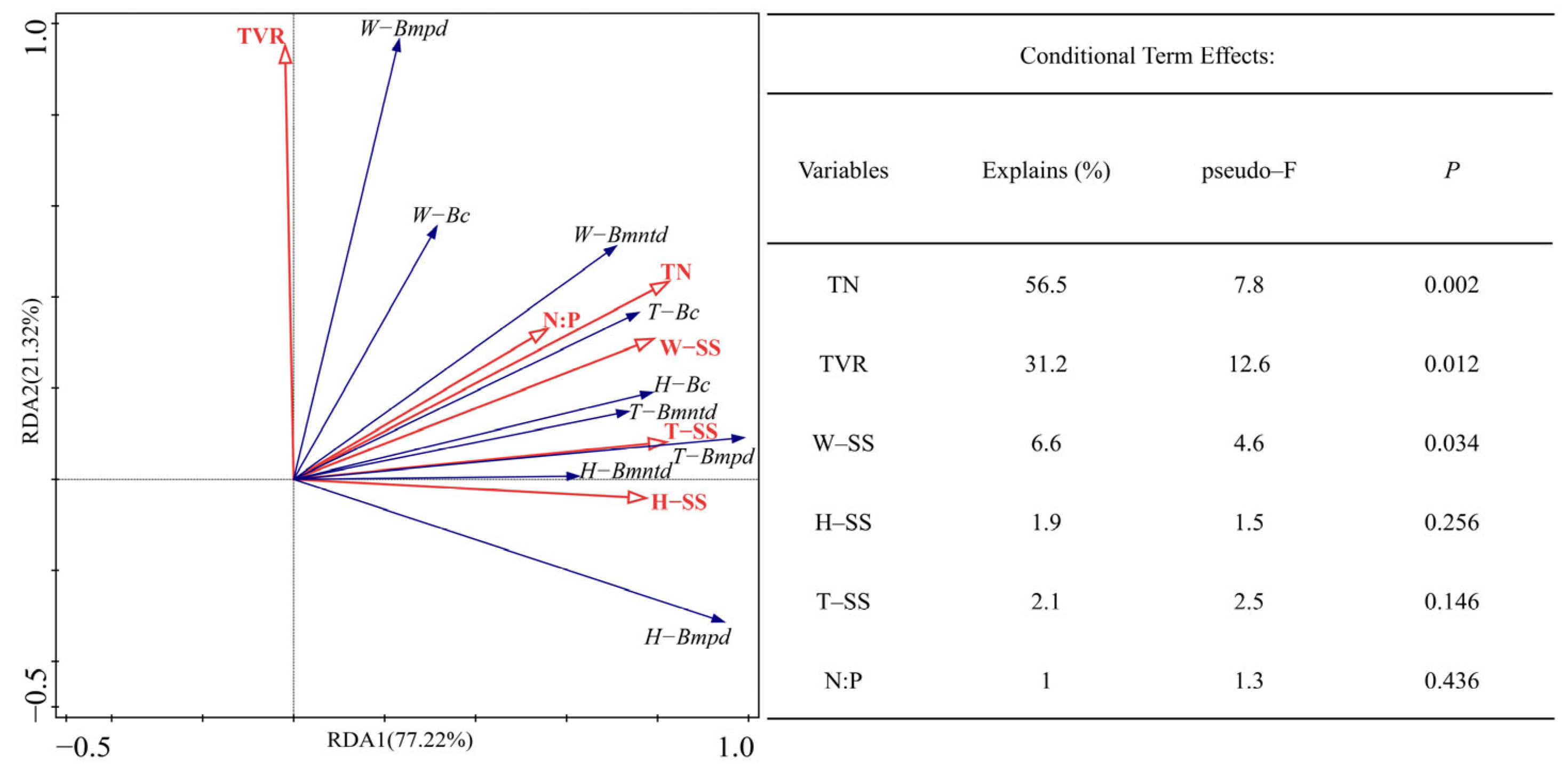{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. The Study Sites and Approach
2.2. Soil Sampling and Laboratory Analyses
2.3. Data Processing
2.4. Statistical Analysis
3. Results and Analysis
3.1. Differences in Vegetation Assemblage between Stages
3.2. Beta Diversity and Endemic Plants in Different Stages
3.3. Patterns of Soil C:N:P Stoichiometry in Different Stages
3.4. Relationship between Beta Diversity and Soil C:N:P Stoichiometry
4. Discussion
4.1. Beta Diversity and Soil C:N:P Stoichiometry in Different Recovery Periods
4.2. Relationships among Beta Diversity, Soil C:N:P Stoichiometry, Time since Vegetation Restoration, and Endemic Plants at Each Stage
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Joly, C.A. The Kunming-Montréal Global Biodiversity Framework. Biota Neotrop. 2023, 22, e2022e001. [Google Scholar] [CrossRef]
- Soto-Navarro, C.A.; Harfoot, M.; Hill, S.L.L.; Campbell, J.; Mora, F.; Campos, C.; Pretorius, C.; Pascual, U.; Kapos, V.; Allison, H. Towards a multidimensional biodiversity index for national application. Nat. Sustain. 2021, 4, 933–942. [Google Scholar] [CrossRef]
- Nicholson, E.; Watermeyer, K.E.; Rowland, J.A.; Sato, C.F.; Stevenson, S.L.; Andrade, A.; Brooks, T.M.; Burgess, N.D.; Cheng, S.-T.; Grantham, H.S.; et al. Scientific foundations for an ecosystem goal, milestones and indicators for the post-2020 global biodiversity framework. Nat. Ecol. Evol. 2021, 5, 1338–1349. [Google Scholar] [CrossRef] [PubMed]
- Isbell, F.; Cowles, J.; Dee, L.E.; Loreau, M.; Reich, P.B.; Gonzalez, A.; Hector, A.; Schmid, B. Quantifying effects of biodiversity on ecosystem functioning across times and places. Ecol. Lett. 2018, 21, 763–778. [Google Scholar] [CrossRef] [Green Version]
- Craven, D.; Eisenhauer, N.; Pearse, W.D.; Hautier, Y.; Isbell, F.; Roscher, C.; Bahn, M.; Beierkuhnlein, C.; Bönisch, G.; Buchmann, N.; et al. Multiple facets of biodiversity drive the diversity–stability relationship. Nat. Ecol. Evol. 2018, 2, 1579–1587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, M.J.; Crist, T.O.; Chase, J.M.; Vellend, M.; Inouye, B.D.; Freestone, A.L.; Sanders, N.J.; Cornell, H.V.; Comita, L.S.; Davies, K.F.; et al. Navigating the multiple meanings of β diversity: A roadmap for the practicing ecologist. Ecol. Lett. 2011, 14, 19–28. [Google Scholar] [CrossRef]
- Le Provost, G.; Schenk, N.V.; Penone, C.; Thiele, J.; Westphal, C.; Allan, E.; Ayasse, M.; Blüthgen, N.; Boeddinghaus, R.S.; Boesing, A.L.; et al. The supply of multiple ecosystem services requires biodiversity across spatial scales. Nat. Ecol. Evol. 2022, 7, 236–249. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, R.H. Vegetation of the Siskiyou mountains, Oregon and California. Ecol. Monogr. 1960, 30, 279–338. [Google Scholar] [CrossRef]
- Condit, R.; Pitman, N.; Leigh, E.G., Jr.; Chave, J.; Terborgh, J.; Foster, R.B.; Nu, P.; Aguilar, S.; Valencia, R.; Hubbell, S.P. Beta-diversity in tropical forest trees. Science 2002, 295, 666–669. [Google Scholar] [CrossRef] [Green Version]
- Socolar, J.B.; Gilroy, J.J.; Kunin, W.E.; Edwards, D.P. How should beta-diversity inform biodiversity conservation? Trends Ecol. Evol. 2016, 31, 67–80. [Google Scholar] [CrossRef] [Green Version]
- Mori, A.S.; Isbell, F.; Seidl, R. β-diversity, community assembly, and ecosystem functioning. Trends Ecol. Evol. 2018, 33, 549–564. [Google Scholar] [CrossRef]
- Reu, J.C.; Catano, C.P.; Spasojevic, M.J.; Myers, J.A. Beta diversity as a driver of forest biomass across spatial scales. Ecology 2022, 103, e3774. [Google Scholar] [CrossRef] [PubMed]
- Weiher, E.; Freund, D.; Bunton, T.; Stefanski, A.; Lee, T.; Bentivenga, S. Advances, challenges and a developing synthesis of ecological community assembly theory. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 2403–2413. [Google Scholar] [CrossRef]
- Myers, J.A.; Chase, J.M.; Jiménez, I.; Jorgensen, P.M.; Araujo-Murakami, A.; Paniagua-Zambrana, N.; Seidel, R. Beta- diversity in temperate and tropical forests reflects dissimilar mechanisms of community assembly. Ecol. Lett. 2013, 16, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Münkemüller, T.; Gallien, L.; Pollock, L.J.; Barros, C.; Carboni, M.; Chalmandrier, L.; Mazel, F.; Mokany, K.; Roquet, C.; Smyčka, J.; et al. Dos and don’ts when inferring assembly rules from diversity patterns. Glob. Ecol. Biogeogr. 2020, 29, 1212–1229. [Google Scholar] [CrossRef] [Green Version]
- Leibold, M.A.; Holyoak, M.; Mouquet, N.; Amarasekare, P.; Chase, J.M.; Hoopes, M.F.; Holt, R.D.; Shurin, J.B.; Law, R.; Tilman, D.; et al. The metacommunity concept: A framework for multi-scale community ecology. Ecol. Lett. 2004, 7, 601–613. [Google Scholar] [CrossRef]
- Kraft, N.J.; Adler, P.B.; Godoy, O.; James, E.C.; Fuller, S.; Levine, J.M. Community assembly, coexistence and the environmental filtering metaphor. Funct. Ecol. 2015, 29, 592–599. [Google Scholar] [CrossRef]
- Chave, J. Neutral theory and community ecology. Ecol. Lett. 2004, 7, 241–253. [Google Scholar] [CrossRef]
- Hubbell, S.P. The unified neutral theory of biodiversity and biogeography (MPB-32). In The Unified Neutral Theory of Biodiversity and Biogeography (MPB-32); Princeton University Press: Princeton, NJ, USA, 2011. [Google Scholar] [CrossRef]
- Chase, J.M.; Myers, J.A. Disentangling the importance of ecological niches from stochastic processes across scales. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 2351–2363. [Google Scholar] [CrossRef] [Green Version]
- Bellier, E.; Grøtan, V.; Engen, S.; Schartau, A.K.; Herfindal, I.; Finstad, A.G. Distance decay of similarity, effects of environmental noise and ecological heterogeneity among species in the spatio-temporal dynamics of a dispersal-limited community. Ecography 2014, 37, 172–182. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Y.; Li, M.; He, N.; Li, J. Divergent roles of environmental and spatial factors in shaping plant β-diversity of different growth forms in drylands. Glob. Ecol. Conserv. 2021, 26, e01487. [Google Scholar] [CrossRef]
- Jiang, L.; Hu, D.; Wang, H.; Lv, G. Discriminating ecological processes affecting different dimensions of α-and β-diversity in desert plant communities. Ecol. Evol. 2022, 12, e8710. [Google Scholar] [CrossRef]
- Wang, Z.; Fang, J.; Tang, Z.; Shi, L. Geographical patterns in the beta diversity of China’s woody plants: The influence of space, environment and range size. Ecography 2012, 35, 1092–1102. [Google Scholar] [CrossRef] [Green Version]
- Fitzpatrick, M.C.; Sanders, N.J.; Normand, S.; Svenning, J.C.; Ferrier, S.; Gove, A.D.; Dunn, R.R. Environmental and historical imprints on beta diversity: Insights from variation in rates of species turnover along gradients. Proc. R. Soc. B Biol. Sci. 2013, 280, 20131201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, S.J.; Salpeter, K.; Comita, L.S. Higher β-diversity observed for herbs over woody plants is driven by stronger habitat filtering in a tropical understory. Ecology 2016, 97, 2074–2084. [Google Scholar] [CrossRef]
- Su, L.; Du, H.; Zeng, F.; Peng, W.; Wang, H.; Wang, K.; Lu, M. Environmental and spatial contributions to tree community assembly across life stages and scales in evergreen-deciduous broadleaf karst forests, southwest China. J. For. Res. 2022, 1–9. [Google Scholar] [CrossRef]
- Hu, D.; Jiang, L.; Hou, Z.; Zhang, J.; Wang, H.; Lv, G. Environmental filtration and dispersal limitation explain different aspects of beta diversity in desert plant communities. Glob. Ecol. Conserv. 2022, 33, e01956. [Google Scholar] [CrossRef]
- Hua, F.; Bruijnzeel, L.A.; Meli, P.; Martin, P.A.; Zhang, J.; Nakagawa, S.; Miao, X.; Wang, W.; Mcevoy, C.; Balmford, A. The biodiversity and ecosystem service contributions and trade-offs of forest restoration approaches. Science 2022, 376, 839–844. [Google Scholar] [CrossRef]
- Pennisi, E. How to regrow a forest? Scientists aren’t sure. Science 2022, 378, 816–817. [Google Scholar] [CrossRef]
- Wu, N.; Qu, Y.; Guse, B.; Makarevičiūtė, K.; To, S.; Riis, T.; Fohrer, N. Hydrological and environmental variables outperform spatial factors in structuring species, trait composition, and beta diversity of pelagic algae. Ecol. Evol. 2018, 8, 2947–2961. [Google Scholar] [CrossRef]
- Qu, Y.; Wu, N.; Guse, B.; Makarevičiūtė, K.; Sun, X.; Fohrer, N. Riverine phytoplankton functional groups response to multiple stressors variously depending on hydrological periods. Ecol. Indic. 2019, 101, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Finderup Nielsen, T.; Sand-Jensen, K.; Dornelas, M.; Bruun, H.H. More is less: Net gain in species richness, but biotic homogenization over 140 years. Ecol. Lett. 2019, 22, 1650–1657. [Google Scholar] [CrossRef]
- Blowes, S.A.; Supp, S.R.; Antão, L.H.; Bates, A.; Bruelheide, H.; Chase, J.M.; Moyes, F.; Magurran, A.; Mcgill, B.; Dornelas, M. The geography of biodiversity change in marine and terrestrial assemblages. Science 2019, 366, 339–345. [Google Scholar] [CrossRef] [Green Version]
- Lindholm, M.; Alahuhta, J.; Heino, J.; Toivonen, H. Temporal beta diversity of lake plants is determined by concomitant changes in environmental factors across decades. J. Ecol. 2021, 109, 819–832. [Google Scholar] [CrossRef]
- Wu, N.; Wang, Y.; Wang, Y.; Sun, X.; Faber, C.; Fohrer, N. Environment regimes play an important role in structuring trait-and taxonomy-based temporal beta diversity of riverine diatoms. J. Ecol. 2022, 110, 1442–1454. [Google Scholar] [CrossRef]
- West, D.C.; Shugart, H.H.; Botkin, D.F. (Eds.) Forest Succession: Concepts and Application; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Sardans, J.; Rivas-Ubach, A.; Peñuelas, J. The C: N: P stoichiometry of organisms and ecosystems in a changing world: A review and perspectives. Perspect. Plant Ecol. Evol. Syst. 2012, 14, 33–47. [Google Scholar] [CrossRef]
- Chen, X.; Chen, H.Y. Plant mixture balances terrestrial ecosystem C: N: P stoichiometry. Nat. Commun. 2021, 12, 4562. [Google Scholar] [CrossRef]
- Lu, J.; Feng, S.; Wang, S.; Zhang, B.; Ning, Z.; Wang, R.; Chen, X.; Yu, L.; Zhao, H.; Lan, D.; et al. Patterns and driving mechanism of soil organic carbon, nitrogen, and phosphorus stoichiometry across northern China’s desert-grassland transition zone. Catena 2023, 220, 106695. [Google Scholar] [CrossRef]
- Güsewell, S. N: P ratios in terrestrial plants: Variation and functional significance. New Phytol. 2004, 164, 243–266. [Google Scholar] [CrossRef]
- He, Y.; Liang, S.; Liu, R.; Jiang, Y. Beta diversity patterns unlock the community assembly of woody plant communities in the riparian zone. Forests 2022, 13, 673. [Google Scholar] [CrossRef]
- Tian, Q.; Zhang, X.; Xu, X.; Yi, H.; He, J.; He, L.; Sun, W. Knowledge about Plant Coexistence during Vegetation Succession for Forest Management on the Loess Plateau, China. Forests 2022, 13, 1456. [Google Scholar] [CrossRef]
- Lohbeck, M.; Poorter, L.; Lebrija-Trejos, E.; Martínez-Ramos, M.; Meave, J.A.; Paz, H.; Pérez-García, E.A.; Romero-Pérez, I.E.; Tauro, A.; Bongers, F. Successional changes in functional composition contrast for dry and wet tropical forest. Ecology 2013, 94, 1211–1216. [Google Scholar] [CrossRef]
- Li, T.; Wu, M.; Duan, C.; Li, S.; Liu, C. The effect of different restoration approaches on vegetation development in metal mines. Sci. Total Environ. 2022, 806, 150626. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Yan, Y.; Zhang, J.; Zhang, Q.; Niu, J. Taxonomic, functional, and phylogenetic beta diversity in the Inner Mongolia grassland. Glob. Ecol. Conserv. 2021, 28, e01634. [Google Scholar] [CrossRef]
- Fang, J.Y.; Zhu, J.L.; Guo, Z.D.; Wang, X.; Shen, Z.; Tang, Z.; He, J.; Yu, D.; Jiang, Y.; Wang, Z.; et al. Methods and protocols for plant community inventory. Biodivers. Sci. 2009, 17, 533–548. [Google Scholar] [CrossRef]
- Blois, J.L.; Williams, J.W.; Fitzpatrick, M.C.; Jackson, S.T.; Ferrier, S. Space can substitute for time in predicting climate-change effects on biodiversity. Proc. Natl. Acad. Sci. USA 2013, 110, 9374–9379. [Google Scholar] [CrossRef] [Green Version]
- Zou, H.Y.; Liu, G.B.; Wang, H.S. The vegetation development in North Ziwuling forest region in last fifty years. Acta Botan. Boreali-Occiden. Sin. 2002, 22, 1–8. (In Chinese) [Google Scholar]
- Fan, W.Y.; Wang, X.A.; Guo, H. Analysis of plant community successional series in the Ziwuling area on the Loess Plateau. Acta Ecol. Sin. 2006, 26, 706–714. (In Chinese) [Google Scholar]
- Deng, L.; Wang, K.B.; Chen, M.L.; Shangguan, Z.P.; Sweeney, S. Soil organic carbon storage capacity positively related to forest succession on the Loess Plateau, China. Catena 2013, 110, 1–7. [Google Scholar] [CrossRef]
- Zhao, Y.G.; Liu, X.F.; Wang, Z.L.; Zhao, S.W. Soil organic carbon fractions and sequestration across a 150-yr secondary forest chronosequence on the Loess Plateau, China. Catena 2015, 133, 303–308. [Google Scholar] [CrossRef]
- Liu, Y.; Zhu, G.; Hai, X.; Li, J.; Shangguan, Z.P.; Peng, C.; Deng, L. Long-term Forest succession improves plant diversity and soil quality but not significantly increase soil microbial diversity: Evidence from the Loess Plateau. Ecol. Eng. 2020, 142, 105631. [Google Scholar] [CrossRef]
- Li, J. Flora of China. Harv. Pap. Bot. 2007, 13, 301–302. [Google Scholar] [CrossRef]
- Haq, S.M.; Tariq, A.; Li, Q.; Yaqoob, U.; Majeed, M.; Hassan, M.; Fatima, S.; Fatima, S.; Bussmann, R.W.; Moazzam, M.F.U.; et al. Influence of Edaphic Properties in Determining Forest Community Patterns of the Zabarwan Mountain Range in the Kashmir Himalayas. Forests 2022, 13, 1214. [Google Scholar] [CrossRef]
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon and organic matter. In Methods of Soil Analysis Part 3—Chemical Methods; American Society of Agronomy, Inc.: Madison, WI, USA, 1996; Volume 5, pp. 961–1010. [Google Scholar] [CrossRef]
- Bremner, J.M. Nitrogen-total. In Methods of Soil Analysis: Part 3 Chemical Methods; American Society of Agronomy, Inc.: Madison, WI, USA, 1996; Volume 5, pp. 1085–1121. [Google Scholar] [CrossRef]
- Schimel, J.P.; Weintraub, M.N. The implications of exoenzyme activity on microbial carbon and nitrogen limitation in soil: A theoretical model. Soil Biol. Biochem. 2003, 35, 549–563. [Google Scholar] [CrossRef]
- Legendre, P.; De Cáceres, M. Beta diversity as the variance of community data: Dissimilarity coefficients and partitioning. Ecol. Lett. 2013, 16, 951–963. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Soly-mos, P.; et al. Vegan: Community Ecology Package. R package version 2.5-6. 2019. Available online: https://CRAN.R-project.org/package=vegan (accessed on 28 May 2021).
- Jin, Y.; Qian, H.V. PhyloMaker2: An updated and enlarged R package that can generate very large phylogenies for vascular plants. Plant Divers. 2022, 44, 335–339. [Google Scholar] [CrossRef]
- Kembel, S.W.; Cowan, P.D.; Helmus, M.R.; Cornwell, W.K.; Morlon, H.; Ackerly, D.D.; Blomberg, S.P.; Webb, C.O. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 2010, 26, 1463–1464. [Google Scholar] [CrossRef] [Green Version]
- Webb, C.O.; Ackerly, D.D.; Kembel, S.W. Phylocom: Software for the analysis of phylogenetic community structure and trait evolution. Bioinformatics 2008, 24, 2098–2100. [Google Scholar] [CrossRef] [Green Version]
- Webb, C.O. Exploring the phylogenetic structure of ecological communities: An example for rain forest trees. Am. Nat. 2000, 156, 145–155. [Google Scholar] [CrossRef]
- Haq, S.M.; Calixto, E.S.; Rashid, I.; Khuroo, A.A. Human-driven disturbances change the vegetation characteristics of temperate forest stands: A case study from Pir Panchal Mountain range in Kashmir Himalaya. Trees For. People 2021, 6, 100134. [Google Scholar] [CrossRef]
- Wu, Z.Y. Distribution type of seed plant genera in China. Kunming: Yunnan Plant Research. Acta Bot. Yunnanica Supp. IV 1991, 1–139. (In Chinese) [Google Scholar]
- Hou, X. 1:1 Million Vegetation Map of China; National Tibetan Plateau Data Center: Beijing, China, 2019. (In Chinese) [Google Scholar]
- Tian, Q.L.; Xu, X.M.; Lyu, D.; Wang, H.J.; Lei, S.Y.; Yi, H.J.; He, J.; He, L.; Xue, F.; Zhou, Y.D.; et al. Relationship between geographical pattern of plant diversity and environmental factors in Beiluo River Basin. China Environ. Sci. 2021, 41, 4378–4387. (In Chinese) [Google Scholar]
- Catano, C.P.; Dickson, T.L.; Myers, J.A. Dispersal and neutral sampling mediate contingent effects of disturbance on plant beta-diversity: A meta-analysis. Ecol. Lett. 2017, 20, 347–356. [Google Scholar] [CrossRef]
- Caccianiga, M.; Luzzaro, A.; Pierce, S.; Ceriani, R.M.; Cerabolini, B. The functional basis of a primary succession resolved by CSR classification. Oikos 2006, 112, 10–20. [Google Scholar] [CrossRef]
- Bazzaz, F.A. Plants in Changing Environments: Linking Physiological, Population, and Community Ecology; Cambridge University Press: Cambridge, UK, 1996. [Google Scholar]
- Abbas, S.; Nichol, J.E.; Zhang, J.; Fischer, G.A. The accumulation of species and recovery of species composition along a 70-year succession in a tropical secondary forest. Ecol. Indic. 2019, 106, 105524. [Google Scholar] [CrossRef]
- Hanusch, M.; He, X.; Ruiz-Hernández, V.; Junker, R.R. Succession comprises a sequence of threshold-induced community assembly processes towards multidiversity. Commun. Biol. 2022, 5, 424. [Google Scholar] [CrossRef]
- Tang, G.L.; Ren, G.Y. Reanalysis of Surface Air Temperature Change of the Last 100 Years over China. Clim. Environ. Res. 2005, 4, 791–798. (In Chinese) [Google Scholar]
- González-Megías, A.; Menendez, R.; Roy, D.; Brereton, T.; Thomas, C.D. Changes in the composition of British butterfly assemblages over two decades. Glob. Change Biol. 2008, 14, 1464–1474. [Google Scholar] [CrossRef]
- Dos Santos, A.S.; Saraiva, D.D.; Müller, S.C.; Overbeck, G.E. Interactive effects of environmental filtering predict beta-diversity patterns in a subtropical forest metacommunity. Perspect. Plant Ecol. Evol. Syst. 2015, 17, 96–106. [Google Scholar] [CrossRef]
- Hamid, M.; Khuroo, A.A.; Charles, B.; Ahmad, R.; Singh, C.P.; Aravind, N.A. Impact of climate change on the distribution range and niche dynamics of Himalayan birch, a typical treeline species in Himalayas. Biodivers. Conserv. 2019, 28, 2345–2370. [Google Scholar] [CrossRef]
- Liang, M.; Baiser, B.; Hallett, L.M.; Hautier, Y.; Jiang, L.; Loreau, M.; Record, S.; Sokol, E.R.; Zarnetske, P.L. Consistent stabilizing effects of plant diversity across spatial scales and climatic gradients. Nat. Ecol. Evol. 2022, 6, 1669–1675. [Google Scholar] [CrossRef] [PubMed]
- Levine, J.M.; Bascompte, J.; Adler, P.B.; Allesina, S. Beyond pairwise mechanisms of species coexistence in complex communities. Nature 2017, 546, 56–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chai, Y.; Yue, M.; Liu, X.; Guo, Y.; Wang, M.; Xu, J.; Zhang, C.; Chen, Y.; Zhang, L. Patterns of taxonomic, phylogenetic diversity during a long-term succession of forest on the Loess Plateau, China: Insights into assembly process. Sci. Rep. 2016, 6, 27087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, D.; He, F. Environmental filtering explains a U-shape latitudinal pattern in regional β-deviation for eastern North American trees. Ecol. Lett. 2019, 22, 284–291. [Google Scholar] [CrossRef]
- Tao, Y.; Zhou, X.B.; Zhang, S.H.; Lu, H.Y.; Shao, H. Soil nutrient stoichiometry on linear sand dunes from a temperate desert in Central Asia. Catena 2020, 195, 104847. [Google Scholar] [CrossRef]
- Feng, J.; He, K.; Zhang, Q.; Han, M.; Zhu, B. Changes in plant inputs alter soil carbon and microbial communities in forest ecosystems. Glob. Change Biol. 2022, 28, 3426–3440. [Google Scholar] [CrossRef]
- Wang, Z.F.; Zheng, F.L. C, N, and P stoichiometric characteristics of Pinus tabulaeformis plantation in the Ziwuling Region of the Loess Plateau. Acta Ecol. Sin. 2018, 38, 6870–6880. [Google Scholar] [CrossRef]
- Chen, X.; Feng, J.; Ding, Z.; Tang, M.; Zhu, B. Changes in soil total, microbial and enzymatic CNP contents and stoichiometry with depth and latitude in forest ecosystems. Sci. Total Environ. 2022, 816, 151583. [Google Scholar] [CrossRef]
- Tang, B.; Rocci, K.S.; Lehmann, A.; Rillig, M.C. Nitrogen increases soil organic carbon accrual and alters its functionality. Glob. Change Biol. 2023, 25, 2325–2337. [Google Scholar] [CrossRef]
- Niu, S.; Song, L.; Wang, J.; Luo, Y.; Yu, G. Dynamic carbon-nitrogen coupling under global change. Sci. China Life Sci. 2023, 1–12. [Google Scholar] [CrossRef]
- Guiz, J.; Ebeling, A.; Eisenhauer, N.; Hacker, N.; Hertzog, L.; Oelmann, Y.; Roscher, C.; Wagg, C.; Hillebrand, H. Interspecific competition alters leaf stoichiometry in 20 grassland species. Oikos 2018, 127, 903–914. [Google Scholar] [CrossRef]
- Finegan, B. Forest succession. Nature 1984, 312, 109–114. [Google Scholar] [CrossRef]
- Furey, G.N.; Tilman, D. Plant biodiversity and the regeneration of soil fertility. Proc. Natl. Acad. Sci. USA 2021, 118, e2111321118. [Google Scholar] [CrossRef]
- Huang, L.; Hu, H.; Bao, W.; Hu, B.; Liu, J.; Li, F. Shifting soil nutrient stoichiometry with soil of variable rock fragment contents and different vegetation types. Catena 2023, 220, 106717. [Google Scholar] [CrossRef]
- Weidlich, E.W.; Von Gillhaussen, P.; Delory, B.M.; Blossfeld, S.; Poorter, H.; Temperton, V.M. The importance of being first: Exploring priority and diversity effects in a grassland field experiment. Front. Plant Sci. 2017, 7, 2008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grainger, T.N.; Letten, A.D.; Gilbert, B.; Fukami, T. Applying modern coexistence theory to priority effects. Proc. Natl. Acad. Sci. USA 2019, 116, 6205–6210. [Google Scholar] [CrossRef] [Green Version]
- Cattelino, P.J.; Noble, I.R.; Slatyer, R.O.; Kessell, S.R. Predicting the multiple pathways of plant succession. Environ. Manag. 1979, 3, 41–50. [Google Scholar] [CrossRef]
- Villa, P.M.; Martins, S.V.; Diniz, É.S.; de Oliveira Neto, S.N.; Neri, A.V.; Pinto, H., Jr.; Nunes, J.A.; Bueno, M.L.; Ali, A. Taxonomic and functional beta diversity of woody communities along Amazon Forest succession: The relative importance of stand age, soil properties and spatial factor. For. Ecol. Manag. 2021, 482, 118885. [Google Scholar] [CrossRef]
- Zhang, M.; Liu, B.; Li, G.; Kuang, Y.; Yue, X.; Jiang, S.; Liu, J.; Wang, L. The relative and combined effects of herbivore assemblage and soil nitrogen on plant diversity. Sci. China Life Sci. 2022, 65, 830–837. [Google Scholar] [CrossRef]
- Rosindell, J.; Hubbell, S.P.; He, F.; Harmon, L.J.; Etienne, R.S. The case for ecological neutral theory. Trends Ecol. Evol. 2012, 27, 203–208. [Google Scholar] [CrossRef]
- Yan, X.; Levine, J.M.; Kandlikar, G.S. A quantitative synthesis of soil microbial effects on plant species coexistence. Proc. Natl. Acad. Sci. USA 2022, 119, e2122088119. [Google Scholar] [CrossRef] [PubMed]
- Zobel, K. On the species-pool hypothesis and on the quasi-neutral concept of plant community diversity. Folia Geobot. 2001, 36, 3–8. [Google Scholar] [CrossRef]
- Jetz, W.; Rahbek, C.; Colwell, R.K. The coincidence of rarity and richness and the potential signature of history in centres of endemism. Ecol. Lett. 2004, 7, 1180–1191. [Google Scholar] [CrossRef]
- McGill, B.J. Matters of scale. Science 2010, 328, 575–576. [Google Scholar] [CrossRef] [Green Version]
- Yao, S.; Qin, J.; Peng, X.; Zhang, B. The effects of vegetation on restoration of physical stability of a severely degraded soil in China. Ecol. Eng. 2009, 35, 723–734. [Google Scholar] [CrossRef]
- Kou, M. The Characteristics of Erosion-Resistant Plant and Its Community in the Hill and Gully Loess Plateau Region; Research Center of Soil and Water Conservation and Ecological Environment, Chinese Academy of Sciences and Ministry of Education: Beijing, China, 2016. (In Chinese) [Google Scholar]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, Q.; Zhang, X.; Xu, X.; Yi, H.; He, J.; He, L. Beta Diversity of Plant Communities in Relation to Soil C:N:P Stoichiometry across 150 Years of Vegetation Restoration in a Temperate Zone. Forests 2023, 14, 553. https://doi.org/10.3390/f14030553
Tian Q, Zhang X, Xu X, Yi H, He J, He L. Beta Diversity of Plant Communities in Relation to Soil C:N:P Stoichiometry across 150 Years of Vegetation Restoration in a Temperate Zone. Forests. 2023; 14(3):553. https://doi.org/10.3390/f14030553
Chicago/Turabian StyleTian, Qilong, Xiaoping Zhang, Xiaoming Xu, Haijie Yi, Jie He, and Liang He. 2023. "Beta Diversity of Plant Communities in Relation to Soil C:N:P Stoichiometry across 150 Years of Vegetation Restoration in a Temperate Zone" Forests 14, no. 3: 553. https://doi.org/10.3390/f14030553